Deck 19: Population Genetics and Human Evolution
Full screen (f)
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
Question
The Denisovan genome contains sequences that originated from an unknown human species. Using Figure 1, speculate on which species this might be. Is it possible that there are other ancestral species that may remain to be discovered that would change the phylogeny presented in the figure?
FIGURE 1 Estimates for the dates of origin and extinction for the three main groups of hominins ( green , blue , and orange ). The australopithecines split into two groups about 2.7 million years ago. One of those groups, the genus Homo , contains the ancestors to our species, H. sapiens.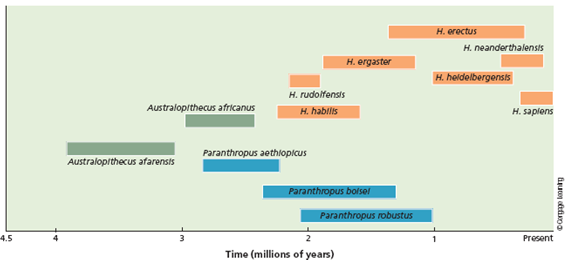
FIGURE 1 Estimates for the dates of origin and extinction for the three main groups of hominins ( green , blue , and orange ). The australopithecines split into two groups about 2.7 million years ago. One of those groups, the genus Homo , contains the ancestors to our species, H. sapiens.
Question
Using Table 1, determine the frequencies of p and q that result in the greatest proportion of heterozygotes in a population.
TablE 1 Heterozygote Frequencies for Recessive Traits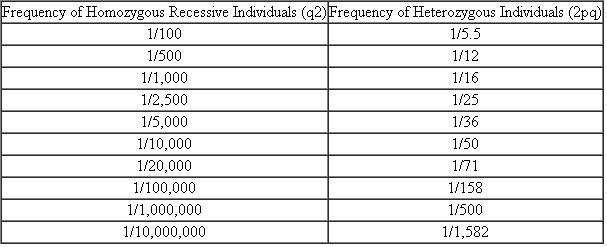
TablE 1 Heterozygote Frequencies for Recessive Traits
Question
Question
Question
Question

Unlock Deck
Sign up to unlock the cards in this deck!
Unlock Deck
Unlock Deck
1/29
Play
Full screen (f)
Deck 19: Population Genetics and Human Evolution
1
Define the following terms:
A) population
B) gene pool
C) allele frequency
D) genotype frequency
A) population
B) gene pool
C) allele frequency
D) genotype frequency
(A) Population - organization of specific species that share a common gene pool.
(b) Gene pool - a set of genetic information that is carried by members of a sexually reproducing population. Genetic information is information that is passed down to the offspring from the parent.
(c) Allele frequency - the occurrence of alleles of a particular gene in a population.
(d) Genotype frequency - pertains to a population. The genotype frequency describes the amount of genotypes in a population. Genotype defines the genetic make-up of an organism.
(b) Gene pool - a set of genetic information that is carried by members of a sexually reproducing population. Genetic information is information that is passed down to the offspring from the parent.
(c) Allele frequency - the occurrence of alleles of a particular gene in a population.
(d) Genotype frequency - pertains to a population. The genotype frequency describes the amount of genotypes in a population. Genotype defines the genetic make-up of an organism.
2
Successful adaptation is defined by:
A) evolving new traits
B) producing many offspring
C) leaving more offspring than others
D) moving to a new location
A) evolving new traits
B) producing many offspring
C) leaving more offspring than others
D) moving to a new location
Successful adaptation is defined by the ability of individuals to produce more offspring than others. So, the rate of reproduction is what matters most in natural selection.
Option (c) leaing more offspring than others is correct.
Although evolving new traits may provide the first step towards adaptation, it may also produce a useless trait that is not advantageous for adaptation. Therefore, option (a) is invalid. Option (b) is almost correct, expect it leaves out that adaptation involves competitiveness in the reproduction rate. Simply having a high number of offspring is not enough for adaptation. The individual must produce offspring at a higher rate then its counterparts for successful adaptation to occur.
Option (d) is invalid because simply moving to a new location does not reflect any type of adaptation. If the organism is able to develop new traits in this new environment, faster than its counterparts, then it is able to adapt. So although moving to a new location is a catalyst for adaptation, it is not defined by it.
Option (c) leaing more offspring than others is correct.
Although evolving new traits may provide the first step towards adaptation, it may also produce a useless trait that is not advantageous for adaptation. Therefore, option (a) is invalid. Option (b) is almost correct, expect it leaves out that adaptation involves competitiveness in the reproduction rate. Simply having a high number of offspring is not enough for adaptation. The individual must produce offspring at a higher rate then its counterparts for successful adaptation to occur.
Option (d) is invalid because simply moving to a new location does not reflect any type of adaptation. If the organism is able to develop new traits in this new environment, faster than its counterparts, then it is able to adapt. So although moving to a new location is a catalyst for adaptation, it is not defined by it.
3
Tracing Ancient Migrations
Genomic information is being used to trace the paths followed by ancient migrations of our species out of Africa to all parts of the Earth. A logical question is: How can we map out events that occurred thousands of years ago that left no written records? The answer is written in the genomes of present-day populations. To work out these routes, geneticists use genetic markers. The markers used in this work are Y chromosome sequences, which are passed directly from father to son, and mitochondrial sequences, which are passed from a mother to all her children. These markers allow men to trace their paternal heritage and men and women to trace their maternal heritage. Because these DNA markers do not undergo recombination during meiosis, mutations that arise in these DNA sequences become heritable markers. These new mutations spread through the population from generation to generation. After many generations, a specific marker will be carried by most members of a population living in a particular geographical region. If people leave that region, they carry that marker with them, and along the way pass it on to their offspring, making its path traceable. The relative ages of markers can be established by assuming that mutations in the markers are random and occur at a constant rate. This assumption is more reliable for Y chromosome markers than for mitochondrial markers but is still useful for establishing the relative ages of each marker.
Ancient migration routes are traced by cataloging the markers present in existing indigenous populations. Knowing the markers characteristic of many indigenous populations provides a starting point from which researchers can work back to track the markers through different populations. DNA samples donated by about 10,000 members of indigenous and traditional peoples from around the world form the starting-point database. Each set of markers we carry represents an ancient point of origin and an end point (where we are now) along a path of migration. By surveying many people in present-day populations, the track of each marker can be reconstructed.
What this means for all of us is that it is now possible to trace our heritage far beyond grandparents and great-grandparents to ancestors who lived thousands of years ago, and to follow the path of their ancient migrations that lead to us and where we live now. The Genographic Project is assembling the largest database for these studies. Part of the database is made up of DNA samples from the 5,000-or-so indigenous populations that have lived in particular regions for many generations and have maintained their languages and cultures. However, the project is also selling kits to those who wish to contribute their DNA, using swabs to collect cheek cells. Online vendors offer similar kits. Others offer autosomal DNA testing using SNPs to provide a large-scale view of someone's heritage, but these tests do not have the specificity of tests using Y chromosome and mitochondrial markers.
Modern forms of H. sapiens spread through central Asia some 50,000 to 70,000 years ago and into Southeast Asia and Australia about 40,000 to 60,000 years ago. H. sapiens moved into Europe some 40,000 to 50,000 years ago, displacing the Neanderthals who had lived there from about 100,000 years ago to about 30,000 years ago.
Genetic data and recent archaeological findings indicate that North America and South America were populated by three or four waves of migration that occurred 15,000 to 30,000 years ago. Migrations from Asia across the Bering Sea are well supported by archaeological and genetic findings, but some researchers feel that Asia may not have been the only source of the first Americans. Some skeletal remains, such as Kennewick Man and the Spirit Cave mummy, have anatomical features that more closely resemble Europeans than Asians. Evidence from mitochondrial DNA haplotype X, found in western Eurasians but not East Asians, and a reinterpretation of stone-tool technology make it seem possible that Europeans migrated to North America more than 10,000 years ago. Some genetic evidence argues against this possibility, but based on the analysis of mitochondrial, Y chromosome, and autosomal DNA from remains excavated in Central Siberia, the results suggest that Native American genomes are of mixed origins, derived from east Asians and western Eurasians.
Further evidence from genetics, anthropology, archeology, and linguistics will be required to provide the final answers about the origins and migrations of the first people to reach the Americas.
Why don't genetic markers on the Y chromosome undergo recombination? Why is this lack of recombination a necessity for these markers to be used in tracing migrations?
Genomic information is being used to trace the paths followed by ancient migrations of our species out of Africa to all parts of the Earth. A logical question is: How can we map out events that occurred thousands of years ago that left no written records? The answer is written in the genomes of present-day populations. To work out these routes, geneticists use genetic markers. The markers used in this work are Y chromosome sequences, which are passed directly from father to son, and mitochondrial sequences, which are passed from a mother to all her children. These markers allow men to trace their paternal heritage and men and women to trace their maternal heritage. Because these DNA markers do not undergo recombination during meiosis, mutations that arise in these DNA sequences become heritable markers. These new mutations spread through the population from generation to generation. After many generations, a specific marker will be carried by most members of a population living in a particular geographical region. If people leave that region, they carry that marker with them, and along the way pass it on to their offspring, making its path traceable. The relative ages of markers can be established by assuming that mutations in the markers are random and occur at a constant rate. This assumption is more reliable for Y chromosome markers than for mitochondrial markers but is still useful for establishing the relative ages of each marker.
Ancient migration routes are traced by cataloging the markers present in existing indigenous populations. Knowing the markers characteristic of many indigenous populations provides a starting point from which researchers can work back to track the markers through different populations. DNA samples donated by about 10,000 members of indigenous and traditional peoples from around the world form the starting-point database. Each set of markers we carry represents an ancient point of origin and an end point (where we are now) along a path of migration. By surveying many people in present-day populations, the track of each marker can be reconstructed.
What this means for all of us is that it is now possible to trace our heritage far beyond grandparents and great-grandparents to ancestors who lived thousands of years ago, and to follow the path of their ancient migrations that lead to us and where we live now. The Genographic Project is assembling the largest database for these studies. Part of the database is made up of DNA samples from the 5,000-or-so indigenous populations that have lived in particular regions for many generations and have maintained their languages and cultures. However, the project is also selling kits to those who wish to contribute their DNA, using swabs to collect cheek cells. Online vendors offer similar kits. Others offer autosomal DNA testing using SNPs to provide a large-scale view of someone's heritage, but these tests do not have the specificity of tests using Y chromosome and mitochondrial markers.
Modern forms of H. sapiens spread through central Asia some 50,000 to 70,000 years ago and into Southeast Asia and Australia about 40,000 to 60,000 years ago. H. sapiens moved into Europe some 40,000 to 50,000 years ago, displacing the Neanderthals who had lived there from about 100,000 years ago to about 30,000 years ago.
Genetic data and recent archaeological findings indicate that North America and South America were populated by three or four waves of migration that occurred 15,000 to 30,000 years ago. Migrations from Asia across the Bering Sea are well supported by archaeological and genetic findings, but some researchers feel that Asia may not have been the only source of the first Americans. Some skeletal remains, such as Kennewick Man and the Spirit Cave mummy, have anatomical features that more closely resemble Europeans than Asians. Evidence from mitochondrial DNA haplotype X, found in western Eurasians but not East Asians, and a reinterpretation of stone-tool technology make it seem possible that Europeans migrated to North America more than 10,000 years ago. Some genetic evidence argues against this possibility, but based on the analysis of mitochondrial, Y chromosome, and autosomal DNA from remains excavated in Central Siberia, the results suggest that Native American genomes are of mixed origins, derived from east Asians and western Eurasians.
Further evidence from genetics, anthropology, archeology, and linguistics will be required to provide the final answers about the origins and migrations of the first people to reach the Americas.
Why don't genetic markers on the Y chromosome undergo recombination? Why is this lack of recombination a necessity for these markers to be used in tracing migrations?
The Y chromosome is one of the two sex chromosomes in mammals. The chromosome contains SRY (Sex determining Region Y) gene whose product stimulates testis development. The Y chromosome is paternally inherited by males only.
During the course of evolution , genes beneficial to males accumulated near the SRY. Any recombination event between X and Y chromosomes resulted in deleterious effects. As a result, recombination in this region was suppressed or prevented to ensure preservation of male specific region. Moreover, genes at SRY are so closely associated that recombination is almost impossible.
Due to these reasons, over time, the Y chromosome evolved to inhibit the region around SRY from recombining with the X chromosome. Only the tips of the Y and X chromosomes are capable of recombination.
Lack of recombination preserves the DNA (Deoxyribo Nucleic Acid) markers in the Y chromosome. The only changes that occur in them are mutations. These mutations become heritable markers when they spread from generation to generation, in a population. The markers will be carried by most of the population living at a region, after certain number of generations.
When people bearing a specific marker (on Y chromosome) migrate to another region, all along the way, they pass it to their offspring. The path of this marker could be traced through chromosomal analysis. The relative ages of the markers is estimated based on the fact that mutations are random and occur at a constant rate.
Thus, lack of recombination of Y chromosome and constant mutation rate of genomes provide accurate information in tracing migrations.
During the course of evolution , genes beneficial to males accumulated near the SRY. Any recombination event between X and Y chromosomes resulted in deleterious effects. As a result, recombination in this region was suppressed or prevented to ensure preservation of male specific region. Moreover, genes at SRY are so closely associated that recombination is almost impossible.
Due to these reasons, over time, the Y chromosome evolved to inhibit the region around SRY from recombining with the X chromosome. Only the tips of the Y and X chromosomes are capable of recombination.
Lack of recombination preserves the DNA (Deoxyribo Nucleic Acid) markers in the Y chromosome. The only changes that occur in them are mutations. These mutations become heritable markers when they spread from generation to generation, in a population. The markers will be carried by most of the population living at a region, after certain number of generations.
When people bearing a specific marker (on Y chromosome) migrate to another region, all along the way, they pass it to their offspring. The path of this marker could be traced through chromosomal analysis. The relative ages of the markers is estimated based on the fact that mutations are random and occur at a constant rate.
Thus, lack of recombination of Y chromosome and constant mutation rate of genomes provide accurate information in tracing migrations.
4
What is the relationship between founder effects and genetic drift?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
5
Natural selection alters genotypic frequencies by increasing or decreasing fitness (i.e., differential fertility or mortality). There are several examples of selection associated with human genetic disorders. Sickle cell anemia and other abnormal hemoglobins are the best examples of selection in humans. Carriers of the sickle and other hemoglobin mutations are more resistant to malaria than is either homozygous class. Therefore, in areas where malaria is endemic, carriers are less likely to die of malaria and will have proportionally more offspring than will homozygotes, thus passing on more genes. Balancing selection may also have influenced carrier frequencies for more "common" recessive diseases, such as cystic fibrosis in Europeans and Tay-Sachs in the Ashkenazi Jewish population, but the selective agent is not known for certain.
Selection may favor homozygotes over heterozygotes, resulting in an unstable polymorphism. One example is selection against heterozygous fetuses when an Rh? mother carries an Rh1 (heterozygous) fetus. This should result in a gradual elimination of the Rh? allele. However, the high frequency of the Rh? allele in so many populations suggests that other, unknown factors may maintain the Rh? allele in human populations.
If allele frequencies in the hemoglobin gene are influenced by sickle cell anemia on the one hand and by resistance to malaria on the other hand, what factors may cause a change in these allele frequencies over time?
Selection may favor homozygotes over heterozygotes, resulting in an unstable polymorphism. One example is selection against heterozygous fetuses when an Rh? mother carries an Rh1 (heterozygous) fetus. This should result in a gradual elimination of the Rh? allele. However, the high frequency of the Rh? allele in so many populations suggests that other, unknown factors may maintain the Rh? allele in human populations.
If allele frequencies in the hemoglobin gene are influenced by sickle cell anemia on the one hand and by resistance to malaria on the other hand, what factors may cause a change in these allele frequencies over time?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
6
How would a drastic reduction in a population's size affect that population's gene pool?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
7
The MN blood group is a single-gene, two-allele system in which each allele is codominant. Why are such codominant alleles ideal for studies of allele frequencies in a population?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
8
The major factor causing deviations from Hardy- Weinberg equilibrium is:
A) selection
B) nonrandom mating
C) mutation
D) migration
E) early death
A) selection
B) nonrandom mating
C) mutation
D) migration
E) early death
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
9
Tracing Ancient Migrations
Genomic information is being used to trace the paths followed by ancient migrations of our species out of Africa to all parts of the Earth. A logical question is: How can we map out events that occurred thousands of years ago that left no written records? The answer is written in the genomes of present-day populations. To work out these routes, geneticists use genetic markers. The markers used in this work are Y chromosome sequences, which are passed directly from father to son, and mitochondrial sequences, which are passed from a mother to all her children. These markers allow men to trace their paternal heritage and men and women to trace their maternal heritage. Because these DNA markers do not undergo recombination during meiosis, mutations that arise in these DNA sequences become heritable markers. These new mutations spread through the population from generation to generation. After many generations, a specific marker will be carried by most members of a population living in a particular geographical region. If people leave that region, they carry that marker with them, and along the way pass it on to their offspring, making its path traceable. The relative ages of markers can be established by assuming that mutations in the markers are random and occur at a constant rate. This assumption is more reliable for Y chromosome markers than for mitochondrial markers but is still useful for establishing the relative ages of each marker.
Ancient migration routes are traced by cataloging the markers present in existing indigenous populations. Knowing the markers characteristic of many indigenous populations provides a starting point from which researchers can work back to track the markers through different populations. DNA samples donated by about 10,000 members of indigenous and traditional peoples from around the world form the starting-point database. Each set of markers we carry represents an ancient point of origin and an end point (where we are now) along a path of migration. By surveying many people in present-day populations, the track of each marker can be reconstructed.
What this means for all of us is that it is now possible to trace our heritage far beyond grandparents and great-grandparents to ancestors who lived thousands of years ago, and to follow the path of their ancient migrations that lead to us and where we live now. The Genographic Project is assembling the largest database for these studies. Part of the database is made up of DNA samples from the 5,000-or-so indigenous populations that have lived in particular regions for many generations and have maintained their languages and cultures. However, the project is also selling kits to those who wish to contribute their DNA, using swabs to collect cheek cells. Online vendors offer similar kits. Others offer autosomal DNA testing using SNPs to provide a large-scale view of someone's heritage, but these tests do not have the specificity of tests using Y chromosome and mitochondrial markers.
Modern forms of H. sapiens spread through central Asia some 50,000 to 70,000 years ago and into Southeast Asia and Australia about 40,000 to 60,000 years ago. H. sapiens moved into Europe some 40,000 to 50,000 years ago, displacing the Neanderthals who had lived there from about 100,000 years ago to about 30,000 years ago.
Genetic data and recent archaeological findings indicate that North America and South America were populated by three or four waves of migration that occurred 15,000 to 30,000 years ago. Migrations from Asia across the Bering Sea are well supported by archaeological and genetic findings, but some researchers feel that Asia may not have been the only source of the first Americans. Some skeletal remains, such as Kennewick Man and the Spirit Cave mummy, have anatomical features that more closely resemble Europeans than Asians. Evidence from mitochondrial DNA haplotype X, found in western Eurasians but not East Asians, and a reinterpretation of stone-tool technology make it seem possible that Europeans migrated to North America more than 10,000 years ago. Some genetic evidence argues against this possibility, but based on the analysis of mitochondrial, Y chromosome, and autosomal DNA from remains excavated in Central Siberia, the results suggest that Native American genomes are of mixed origins, derived from east Asians and western Eurasians.
Further evidence from genetics, anthropology, archeology, and linguistics will be required to provide the final answers about the origins and migrations of the first people to reach the Americas.
The presence of haplotype X in Native American genomes could have originated from gene flow from western Eurasians into Siberians or by migration of early Europeans across the Atlantic Ocean. What kind of evidence would help resolve this issue?
Genomic information is being used to trace the paths followed by ancient migrations of our species out of Africa to all parts of the Earth. A logical question is: How can we map out events that occurred thousands of years ago that left no written records? The answer is written in the genomes of present-day populations. To work out these routes, geneticists use genetic markers. The markers used in this work are Y chromosome sequences, which are passed directly from father to son, and mitochondrial sequences, which are passed from a mother to all her children. These markers allow men to trace their paternal heritage and men and women to trace their maternal heritage. Because these DNA markers do not undergo recombination during meiosis, mutations that arise in these DNA sequences become heritable markers. These new mutations spread through the population from generation to generation. After many generations, a specific marker will be carried by most members of a population living in a particular geographical region. If people leave that region, they carry that marker with them, and along the way pass it on to their offspring, making its path traceable. The relative ages of markers can be established by assuming that mutations in the markers are random and occur at a constant rate. This assumption is more reliable for Y chromosome markers than for mitochondrial markers but is still useful for establishing the relative ages of each marker.
Ancient migration routes are traced by cataloging the markers present in existing indigenous populations. Knowing the markers characteristic of many indigenous populations provides a starting point from which researchers can work back to track the markers through different populations. DNA samples donated by about 10,000 members of indigenous and traditional peoples from around the world form the starting-point database. Each set of markers we carry represents an ancient point of origin and an end point (where we are now) along a path of migration. By surveying many people in present-day populations, the track of each marker can be reconstructed.
What this means for all of us is that it is now possible to trace our heritage far beyond grandparents and great-grandparents to ancestors who lived thousands of years ago, and to follow the path of their ancient migrations that lead to us and where we live now. The Genographic Project is assembling the largest database for these studies. Part of the database is made up of DNA samples from the 5,000-or-so indigenous populations that have lived in particular regions for many generations and have maintained their languages and cultures. However, the project is also selling kits to those who wish to contribute their DNA, using swabs to collect cheek cells. Online vendors offer similar kits. Others offer autosomal DNA testing using SNPs to provide a large-scale view of someone's heritage, but these tests do not have the specificity of tests using Y chromosome and mitochondrial markers.
Modern forms of H. sapiens spread through central Asia some 50,000 to 70,000 years ago and into Southeast Asia and Australia about 40,000 to 60,000 years ago. H. sapiens moved into Europe some 40,000 to 50,000 years ago, displacing the Neanderthals who had lived there from about 100,000 years ago to about 30,000 years ago.
Genetic data and recent archaeological findings indicate that North America and South America were populated by three or four waves of migration that occurred 15,000 to 30,000 years ago. Migrations from Asia across the Bering Sea are well supported by archaeological and genetic findings, but some researchers feel that Asia may not have been the only source of the first Americans. Some skeletal remains, such as Kennewick Man and the Spirit Cave mummy, have anatomical features that more closely resemble Europeans than Asians. Evidence from mitochondrial DNA haplotype X, found in western Eurasians but not East Asians, and a reinterpretation of stone-tool technology make it seem possible that Europeans migrated to North America more than 10,000 years ago. Some genetic evidence argues against this possibility, but based on the analysis of mitochondrial, Y chromosome, and autosomal DNA from remains excavated in Central Siberia, the results suggest that Native American genomes are of mixed origins, derived from east Asians and western Eurasians.
Further evidence from genetics, anthropology, archeology, and linguistics will be required to provide the final answers about the origins and migrations of the first people to reach the Americas.
The presence of haplotype X in Native American genomes could have originated from gene flow from western Eurasians into Siberians or by migration of early Europeans across the Atlantic Ocean. What kind of evidence would help resolve this issue?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
10
A specific mutation in the BRCA1 gene has been estimated to be present in approximately 1% of Ashkenazi Jewish women of Eastern European descent. This specific alteration, 185delAG, is found about three times more often in this ethnic group than the combined frequency of the other 125 mutations found to date. It is believed that the mutation is the result of a founder effect from many centuries ago. Explain the founder principle.
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
11
Explain the connection between changes in population allele frequencies and evolution, and relate this to the observations made by Wallace and Darwin concerning natural selection.
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
12
The theory of natural selection has been summarized popularly as "survival of the fittest." Is this an accurate description of natural selection? Why or why not?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
13
Can populations evolve without changes in allele frequencies?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
14
Will a recessive allele that is lethal in the homozygous condition ever be completely removed from a large population by natural selection?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
15
Design an experiment to determine if a population is evolving.
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
16
Do you think that our species is still evolving, or are we shielded from natural selection by civilization? Is it possible that misapplications of technology will end up exposing our species to more rather than less natural selection (consider the history of antibiotics)?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
17
What are four assumptions of the Hardy-Weinberg law?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
18
Provide a genetic definition of race.
b. Using this definition, can modern humans be divided into races? Why or why not?
b. Using this definition, can modern humans be divided into races? Why or why not?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
19
Drawing on your newly acquired understanding of the Hardy-Weinberg equilibrium law, point out why the following statement is erroneous: "Because most of the people in Sweden have blond hair and blue eyes, the genes for blond hair and blue eyes must be dominant in that population."
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
20
Briefly describe the two major theories discussed in this chapter about the origin of modern humans.
b. Which of these two theories would predict a closer relationship for the various modern human populations?
c. Which of the two theories is best supported by the genetic evidence?
b. Which of these two theories would predict a closer relationship for the various modern human populations?
c. Which of the two theories is best supported by the genetic evidence?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
21
In a population where the females have the allelic frequencies A = 0.35 and a = 0.65 and the frequencies for males are A = 0.1 and a = 0.9, how many generations will it take to reach Hardy-Weinberg equilibrium for both the allelic and the genotypic frequencies? Assume random mating and show the allelic and genotypic frequencies for each generation.
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
22
The human and chimpanzee genomes are 98.8% identical. If this is so, why are the phenotypes of chimps and humans so different?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
23
Suppose you are monitoring the allelic and genotypic frequencies of the MN blood group locus (see Question 2 for a description of the MN blood group) in a small human population. You find that for 1-year-old children, the genotypic frequencies are MM = 0.25, MN = 0.5, and NN = 0.25, whereas the genotypic frequencies for adults are MM = 0.3, MN = 0.4, and NN = 0.3.
a. Compute the M and N allele frequencies for 1-year-olds and adults.
b. Are the allele frequencies in equilibrium in this population?
c. Are the genotypic frequencies in equilibrium?
a. Compute the M and N allele frequencies for 1-year-olds and adults.
b. Are the allele frequencies in equilibrium in this population?
c. Are the genotypic frequencies in equilibrium?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
24
The Denisovan genome contains sequences that originated from an unknown human species. Using Figure 1, speculate on which species this might be. Is it possible that there are other ancestral species that may remain to be discovered that would change the phylogeny presented in the figure?
FIGURE 1 Estimates for the dates of origin and extinction for the three main groups of hominins ( green , blue , and orange ). The australopithecines split into two groups about 2.7 million years ago. One of those groups, the genus Homo , contains the ancestors to our species, H. sapiens.
FIGURE 1 Estimates for the dates of origin and extinction for the three main groups of hominins ( green , blue , and orange ). The australopithecines split into two groups about 2.7 million years ago. One of those groups, the genus Homo , contains the ancestors to our species, H. sapiens.
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
25
Using Table 1, determine the frequencies of p and q that result in the greatest proportion of heterozygotes in a population.
TablE 1 Heterozygote Frequencies for Recessive Traits
TablE 1 Heterozygote Frequencies for Recessive Traits
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
26
In a given population, the frequencies of the four phenotypic classes of the ABO blood groups are found to be A = 0.33, B = 0.33, AB = 0.18, and i = 0.16. What is the frequency of the i allele?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
27
If a trait determined by an autosomal recessive allele occurs at a frequency of 0.25 in a population, what are the allelic frequencies? Assume Hardy-Weinberg equilibrium and use A and a to symbolize the dominant and recessive alleles, respectively.
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
28
Natural selection alters genotypic frequencies by increasing or decreasing fitness (i.e., differential fertility or mortality). There are several examples of selection associated with human genetic disorders. Sickle cell anemia and other abnormal hemoglobins are the best examples of selection in humans. Carriers of the sickle and other hemoglobin mutations are more resistant to malaria than is either homozygous class. Therefore, in areas where malaria is endemic, carriers are less likely to die of malaria and will have proportionally more offspring than will homozygotes, thus passing on more genes. Balancing selection may also have influenced carrier frequencies for more "common" recessive diseases, such as cystic fibrosis in Europeans and Tay-Sachs in the Ashkenazi Jewish population, but the selective agent is not known for certain.
Selection may favor homozygotes over heterozygotes, resulting in an unstable polymorphism. One example is selection against heterozygous fetuses when an Rh? mother carries an Rh1 (heterozygous) fetus. This should result in a gradual elimination of the Rh? allele. However, the high frequency of the Rh? allele in so many populations suggests that other, unknown factors may maintain the Rh? allele in human populations.
If you suspected that heterozygous carriers of a particular disease gene had a selective advantage in resisting a type of infection, how would you go about testing that hypothesis?
Selection may favor homozygotes over heterozygotes, resulting in an unstable polymorphism. One example is selection against heterozygous fetuses when an Rh? mother carries an Rh1 (heterozygous) fetus. This should result in a gradual elimination of the Rh? allele. However, the high frequency of the Rh? allele in so many populations suggests that other, unknown factors may maintain the Rh? allele in human populations.
If you suspected that heterozygous carriers of a particular disease gene had a selective advantage in resisting a type of infection, how would you go about testing that hypothesis?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
29
Why is it that mutation, acting alone, has little effect on gene frequency?
Unlock Deck
Unlock for access to all 29 flashcards in this deck.
Unlock Deck
k this deck
Unlock Deck
Unlock for access to all 29 flashcards in this deck.


